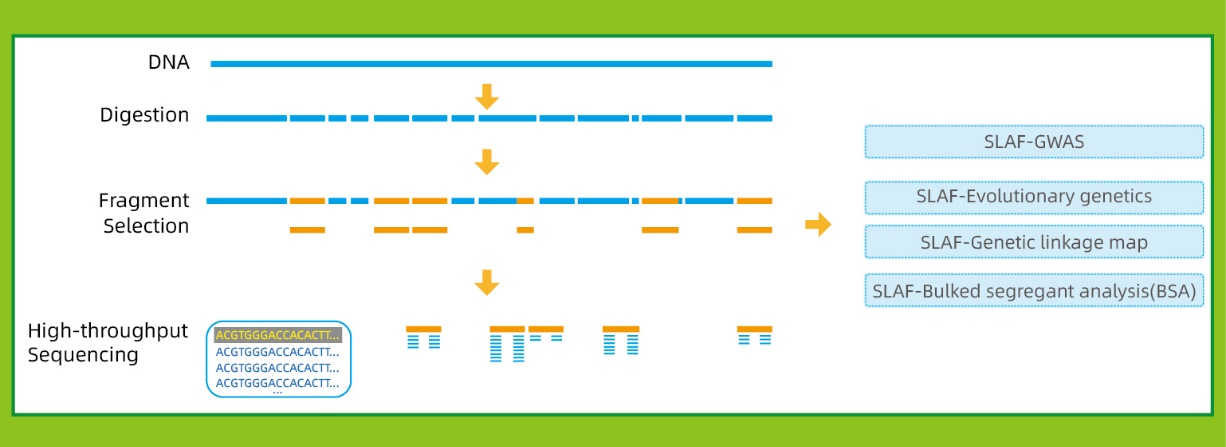
Специфично-локус засилен секвенционирање на фрагменти (SLAF-Seq)

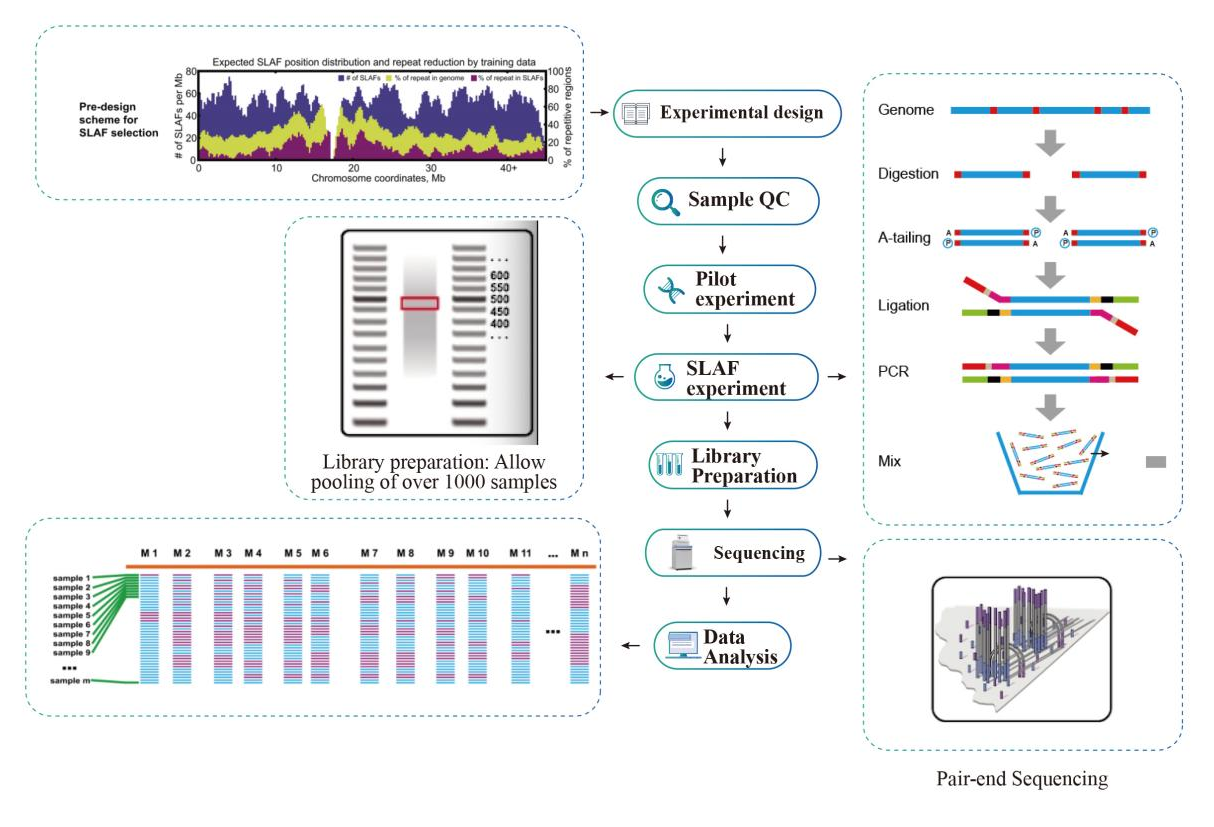
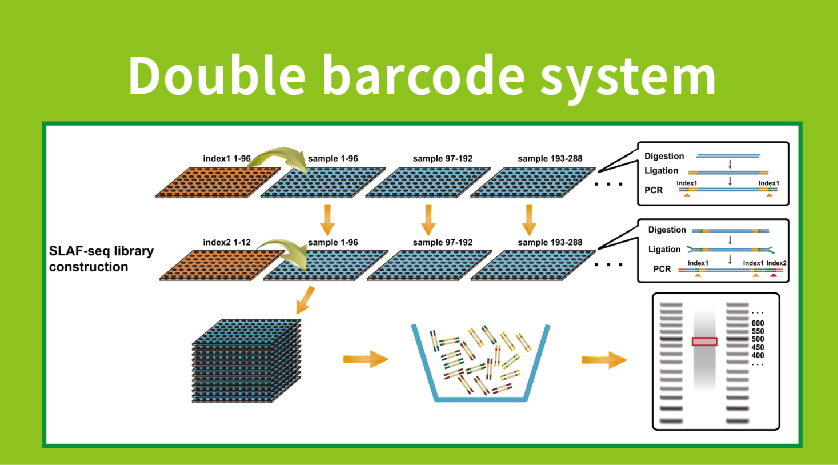
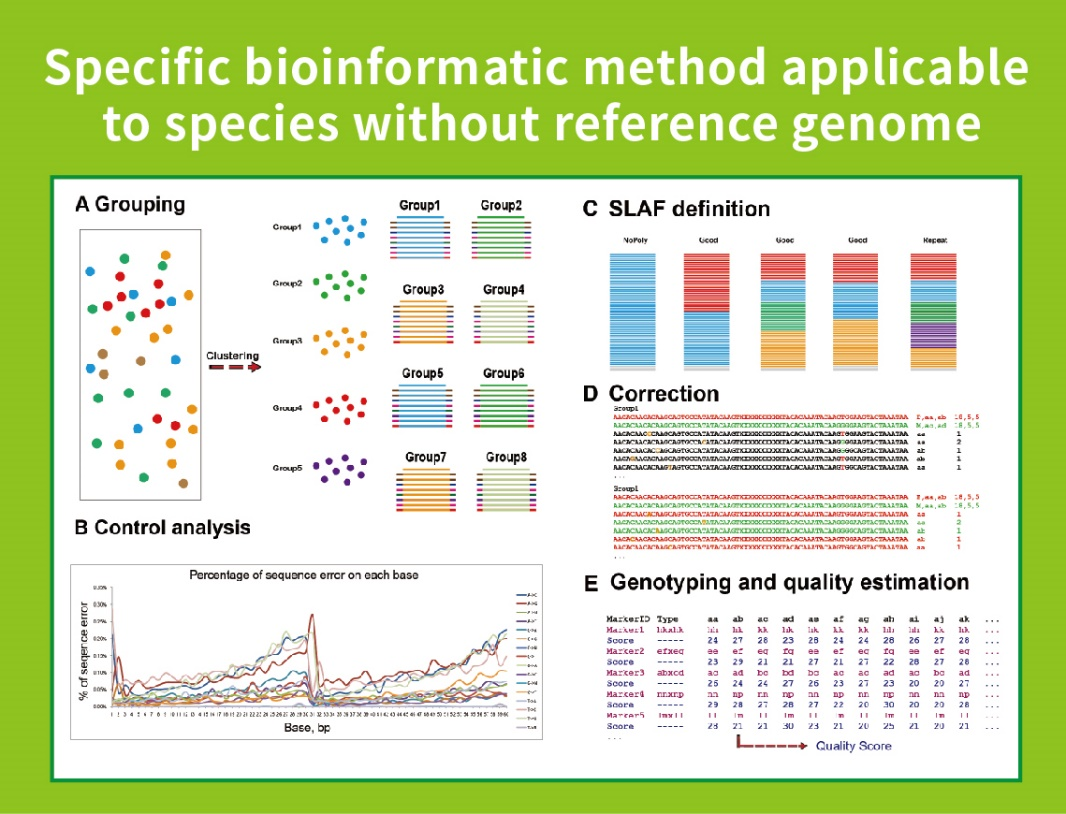
Работен тек

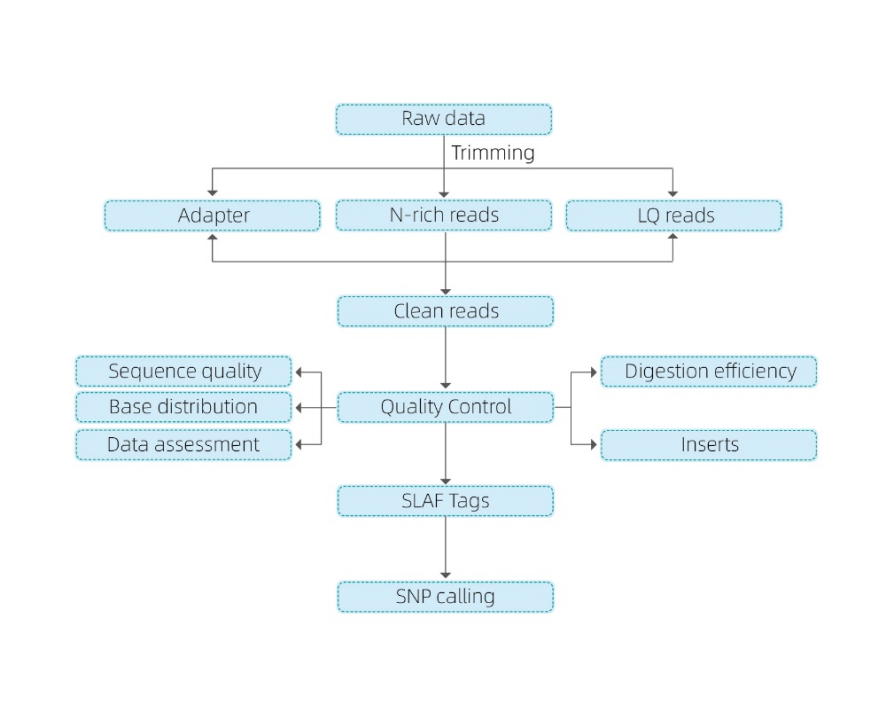
Техничка шема

Карактеристики на услугите
● Секвенционирање на Новасек со PE150.
● Подготовка на библиотеката со двојно баркодирање, овозможувајќи здружување на над 1000 примероци.
● Оваа техника може да се користи со или без референтен геном, со различни биоинформатички цевководи за секој случај:
Со референтен геном: откривање на SNP и Indel
Без референтен геном: кластерирање на примероци и откривање на СНП
● Вово силикоСе прикажуваат повеќекратни комбинации на ензимски фази на фаза на дизајн, за да се најдат оние што генерираат униформа дистрибуција на ознаките SLAF по геномот.
● За време на пред-експериментот, три комбинации на ензими се тестираат во 3 примероци за да се генерираат 9 библиотеки на SLAF, а овие информации се користат за да се избере оптимална комбинација на ензим за ограничување за проектот.
Предности на услугата
●Високо откритие за генетски маркер: Интегрирањето на систем за двојно баркод со голема моќност овозможува истовремено секвенционирање на големите популации, а засилувањето специфично за локус ја подобрува ефикасноста, осигурувајќи дека броевите на ознаките ги исполнуваат различните барања на различни истражувачки прашања.
● Мала зависност од геномот: Може да се примени на видови со или без референтен геном.
●Флексибилен дизајн на шема: Едно-ензим, двоен ензим, мулти-ензимски варење и разни видови на ензими, сите можат да бидат избрани за да се грижат за различни истражувачки цели или видови. Наво силикоПред-дизајнот се спроведува за да се обезбеди оптимален дизајн на ензими.
● Висока ефикасност во ензимското варење: Спроведувањето на анво силикоПред-дизајнот и претходно експериментацијата обезбедени оптимален дизајн со рамномерна дистрибуција на ознаките SLAF на хромозомот (1 SLAF TAG/4KB) и намалена повторлива секвенца (<5%).
●Широка експертиза: Нашиот тим носи богатство на искуство на секој проект, со евиденција за затворање на над 5000 проекти SLAF-Seq на стотици видови, вклучувајќи растенија, цицачи, птици, инсекти и водни организми.
● Само-развиен биоинформатички проток на работа: BMKGene разви интегриран биоинформатички работен тек за SLAF-Seq за да се обезбеди веродостојност и точност на конечниот резултат.
Спецификации на услугите
| Вид на анализа | Препорачана скала на население | Стратегија за секвенцирање | |
| Длабочина на секвенционирање на ознаки | Број на ознака | ||
| Генетски мапи | 2 родители и> 150 потомци | Родители: 20x WGS Offsping: 10x | Големина на геномот: <400 MB: се препорачува WGS <1 GB: 100K ознаки 1-2 GB :: 200K ознаки > 2 GB: 300K ознаки Макс 500к ознаки |
| Студии за асоцијација ширум геном (GWAS) | ≥200 примероци | 10x | |
| Генетска еволуција | ≥ 30 примероци, со> 10 примероци од секоја подгрупа | 10x | |
Барања за услуги
Концентрација ≥ 5 ng/μL
Вкупна количина ≥ 80 ng
Nanodrop OD260/280 = 1,6-2.5
Агарозен гел: Не или ограничена деградација или загадување
Препорачана испорака на примероци
Контејнер: 2 ml центрифуга цевка
(За повеќето примероци, препорачуваме да не зачувате во етанол)
Етикетирање на примерокот: Примероците треба да бидат јасно обележани и идентични за доставениот формулар за информации за примерокот.
Испорака: Сув мраз: Примероците прво треба да бидат спакувани во торби и да се закопаат во сув мраз.
Услужен проток на работа


Пример QC
Пилот експеримент
SLAF-експеримент
Подготовка на библиотека
Секвенцирање
Анализа на податоците
Услуги по продажба
 Вклучува следнава анализа:
Вклучува следнава анализа:

- Податоци за секвенцирање QC
- Развој на SLAF TAG
Мапирање на референтен геном
Без референтен геном: кластерирање
- Анализа на ознаките SLAF: Статистика, дистрибуција преку геномот
- Откривање на маркер: SNP, Indel, SNV, CV Calling и прибелешка
Дистрибуција на ознаки SLAF на хромозомите:

Дистрибуција на СНП на хромозомите:
 Прибелешка на СНП
Прибелешка на СНП

| Година | Journalурнал | IF | Наслов | Апликации |
| 2022 година | Природни комуникации | 17.694 | Геномска основа на гига-хромозомите и гига-геномот на божур од дрво Paeonia ostii | SLAF-GWAS |
| 2015 година | Нов фитолог | 7.433 | Стапалки за припишување прицврстувачки геномски региони од агрономско значење во Соја | SLAF-GWAS |
| 2022 година | Весник на напредно истражување | 12.822 | Вештачки интрогресии ширум геном на Gossypium barbadense во G. hirsutum Откријте супериорни локации за истовремено подобрување на квалитетот и приносот на памучните влакна Карактеристики | SLAF-еволутивна генетика |
| 2019 година | Молекуларно растение | 10.81 | Геномска анализа на населението и собранието на де ново го откриваат потеклото на плевелот Рајс како еволутивна игра | SLAF-еволутивна генетика |
| 2019 година | Генетика на природата | 31.616 | Секвенца на геном и генетска разновидност на обичниот крап, cyprinus carpio | Мапа на Slaf-Linkage |
| 2014 година | Генетика на природата | 25.455 | Геномот на култивиран кикирики дава увид во мешунките кариотипи, полиплоид Еволуција и припишување на земјоделски култури. | Мапа на Slaf-Linkage |
| 2022 година | Весник на растителни биотехнологија | 9.803 | Идентификувањето на ST1 открива селекција што вклучува метеж на морфологија на семето и содржина на нафта за време на припитомување на соја | Развој на SLAF-ознаки |
| 2022 година | Меѓународен весник на молекуларни науки | 6.208 | Идентификација и развој на ДНК маркер за пченица-леимус Молис 2NS (2Д) Замена на хромозомот на дизом | Развој на SLAF-ознаки |
| Година | Journalурнал | IF | Наслов | Апликации |
| 2023 година | Граници во растителна наука | 6.735 | QTL мапирање и транскрипторна анализа на содржината на шеќер за време на овошје со зреење на пирус пирифолија | Генетска мапа |
| 2022 година | Весник на растителни биотехнологија | 8.154 | Идентификувањето на ST1 открива селекција што вклучува метеж на морфологија на семето и содржина на нафта за време на припитомување на соја
| Повикување на СНП |
| 2022 година | Граници во растителна наука | 6.623 | Мапирање на асоцијација ширум геном на Хуллеј едвај фенотипови во околината на сушата.
| Gwas |